La reproducción sexual y las Leyes de Mendel: meiosis y teoría cromosómica de la herencia
La biología celular y molecular moderna ha logrado explicar la base material, y los mecanismos y procesos que permiten demostrar porqué las leyes de Mendel son, efectivamente, leyes científicas. Y es que la genial intuición mendeliana de que los progenitores pasan a sus descendientes una copia de cada factor hereditario (gen) de las dos que poseen, es un hecho comprobado: meiosis es el nombre que recibe el proceso por el que se reparten equitativamente y al azar los genes entre los gametos, de forma que cada gameto recibe una sola copia de cada gen.
Visto desde la perspectiva del presente, es normal afirmar que Mendel fue afortunado al elegir siete rasgos que se trasmiten independientemente unos de otros según la ley de la combinación: hoy conocemos la materia que constituye los genes (el ADN), cómo se empaqueta y cómo se trasmite a los gametos. A diferencia de lo que Mendel propuso, los genes no van en unidades individuales separadas, sino que son fragmentos más o menos grandes de ADN que se integran en gran número en una macromolécula que recibe el nombre de cromosoma. Y es una copia de cada cromosoma lo que se reparte entre los gametos, de forma que muchos genes van a ir juntos en el reparto: es tanto como asegurar que los genes responsables de los siete rasgos utilizados por Mendel tienen por fuerza que hallarse en cromosomas diferentes y, dado que Pisum sativum sólo tiene 7 cromosomas diferentes…
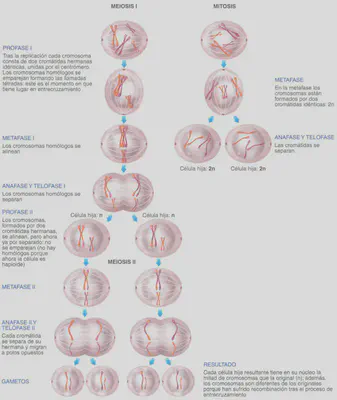
Pero vayamos por partes: la herencia genética ha de ser estrictamente equitativa. Los gametos han de recibir una copia, y sólo una, de cada cromosoma (Fig. 2.7), de forma que cada gameto reciba una copia de cada gen, de las dos que normalmente poseen las células de los organismos pluricelulares. Aquí los conceptos esenciales son diploide y haploide. Las células somáticas normales son diploides, ya que portan dos copias de cada cromosoma (y por ende, de cada gen). Podemos decir que los cromosomas van por parejas en el sentido de que contienen los mismos genes: cada cromosoma de cada una de esas parejas tiene su homólogo. El número diploide de cromosomas se representa con la expresión 2n. Puesto que los gametos son haploides porque sólo contienen una copia de cada cromosoma, su número se representa con n. Los gametos de los guisantes de Mendel, por ejemplo, poseen 7 cromosomas (n=7) y sus células somáticas 14 cromosomas (2n); la especie humana es también diploide y el número normal de cromosomas de nuestras células es de 46 (2n=46); los óvulos y espermatozoides humanos, por su parte, contienen sólo 23 cromosomas (n=23).
El desarrollo de cualquier organismo vivo pluricelular implica reproducción celular: mediante mitosis, las células se multiplican de forma que una célula se divide y da lugar a dos células idénticas: esto implica que las células somáticas humanas que se dividen han de hacer previamente dos copias exactas de todo su material genético y luego, distribuirlo exactamente entre las dos células resultantes de la mitosis. Esto significa que han de sintetizar una copia nueva de cada cromosoma, de forma que cada una de las dos células hijas resultantes tenga a su vez 2n cromosomas, o sea, 46 cromosomas.
En este momento es pertinente delimitar muy bien el concepto de cromosoma en el contexto de la división celular, sea mitótica, sea meiótica. Tanto en la mitosis como en la meiosis 1 (ver Fig. 2.7), al menos hasta la metafase, los cromosomas están formados por dos cromátidas, puesto que la molécula de ADN original ha de haberse replicado, de forma que en este momento del proceso, las células tienen un número diploide de cromosomas formados cada uno por dos cromátidas; seguramente por eso en algunos textos a esta forma del cromosoma lo llaman cromosoma replicado. La esencial diferencia entre mitosis y meiosis 1 es que, al dividirse en dos la célula original, en el caso de la mitosis las células hijas reciben cada una una cromátida de cada cromosoma (con lo que se mantiene el número diploide, sólo que ahora se habla de cromátidas como sinónimo de cromosoma), mientras que en ei caso de la meiosis 1, las dos células resultantes reciben cada una un número haploide de cromosomas (la mitad de los cromosomas), pero todavía constituidos por dos cromátidas cada uno: si en la mitosis humana las dos células resultantes de la división celular contienen 46 cromosomas formados por una sola cromátida cada uno, en el caso de la meiosis 1, el resultado son dos células con 23 cromosomas cada una, pero en este caso formados por dos cromátidas.
La Meiosis Propiamente Dicha
Describiendo el proceso de meiosis se podrá comprender mejor el proceso de trasmisión de la herencia genética. Como se verá, la clave de la vida es la reproducción. La reproducción consiste en hacer copias de sí mismo. Los organismos unicelulares se limitan a dividirse, de forma que de una célula original salen dos, y así sucesivamente. Pero los pluricelulares no tienen esta posibilidad: de hecho, éstos parten de una única célula original, llamada zigoto, que por divisiones sucesivas (mitosis) y diversos procesos de especialización, terminan dando lugar a tales organismos complejos. Puesto que en la reproducción sexual cada progenitor aporta una sola copia de cada gen, es preciso un mecanismo que explique cómo es posible que se formen gametos -que al unirse en la fecundación forman el zigoto- con una sola copia de todos y cada uno de los genes.
La palabra meiosis deriva de un verbo griego que significa reducir, así que hemos de definir la meiosis como división celular reductora. Y ello por una buena razón, porque a partir de una célula diploide (2n cromosomas) hemos de conseguir por división, células haploides de n cromosomas. Pensemos además que si no tuviera lugar esta reducción, en las especies sexuales iría aumentando el número de cromosomas de una manera exponencial, si cada cigoto ha de incorporar el total de cromosomas de cada progenitor. Por tanto, lo esencial para nosotros es entender el proceso por el cual se logra formar gametos haploides a partir de precursores diploides.
La meiosis se lleva a cabo en dos etapas. La primera consiste en dividir la célula (2n) de tal forma que cada célula hija reciba un único y completo juego de cromosomas de la célula madre, es decir, pase a ser haploide (n). Cada célula hija recibe un miembro de cada una de las parejas de cromosomas, sólo uno de los cromosomas homólogos. Esta etapa recibe el nombre de meiosis 1. La segunda fase, denominada meiosis 11, consiste en una división normal, equivalente a una mitosis, de las células obtenidas en la primera etapa (Fig. 2.7).
La meiosis 1 consta de las mismas partes que la mitosis, aunque no son equivalentes. Comienza con la profase I. Ésta difiere de la profase mitótica en un hecho muy importante: los cromosomas homólogos se aparean dos a dos, punto por punto, a lo largo de toda su longitud, formando lo que se denominan bivalentes (por estar compuestos por dos cromosomas homólogos). También reciben el nombre de tétrada, en referencia a las cuatro cromátidas del bivalente; dos por cada cromosoma homólogo (los cromosomas al entrar en la meiosis, como ocurre en la mitosis, están duplicados y por eso constan de dos cromátidas cada uno) (Figs. 2.7 y 2.8).
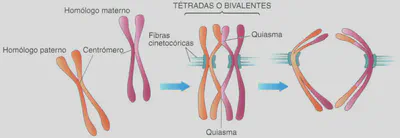
El apareamiento de los homólogos tiene una importancia extraordinaria. A través de él se produce el fenómeno citológico del entrecruzamiento, mediante el cual se lleva a cabo la recombinación génica, el intercambio de genes de un cromosoma homólogo a otro, como luego veremos. La siguiente etapa es la metafase 1. En ella los bivalentes o tétradas (Fig. 2.8), mediante sus centrómeros, se insertan en las fibras del huso adoptando una ordenación circular sobre la placa ecuatorial. La metafase 1 continúa con la anafase 1 en la que, a diferencia de la anafase de una mitosis normal en la que se separan 2n cromátidas, en ésta se separan los cromosomas de los bivalentes, emigrando n cromosomas (cada uno con sus dos cromátidas) a cada polo.
Finalmente, en la telofase 1 los cromosomas se sitúan en ambos polos de la célula, se desespiralizan y se produce la citocinesis, dando lugar a dos células hijas con n cromosomas. Por haberse reducido el número de cromosomas a la mitad, a esta división meiótica se le denomina también división reduccional.
Como se ha indicado, los cromosomas no han seguido durante la meiosis 1 el mismo comportamiento que muestran durante la mitosis, ya que aquí se han separado cromosomas homólogos y no cromátidas. Este movimiento cromosómico es la demostración citológica de la ley de la segregación. Los cromosomas homólogos portan los alelos de cada gen, y su separación es también la separación de esos alelos descrita por Mendel. Las células hijas obtenidas en la meiosis 1 pueden entrar inmediatamente en la meiosis II o pasar por un período de interfase previamente. En cualquier caso, antes de entrar en esta segunda división meiótica, no se produce duplicación de cromosomas, pues ya están duplicados, dado que en la meiosis 1 se separaron cromosomas con sus dos cromátidas. La meiosis II es prácticamente igual que la mitosis, salvo por el hecho de que la célula que entra en división es haploide, ya no hay cromosomas homólogos y, por tanto, tras ella se obtienen dos células hijas con n cromátidas.
Recombinación y Ligamiento
En la profase I se efectúa el emparejamiento de los cromosomas homólogos dos a dos formando las llamadas tétradas o bivalentes (Fig. 2.8). Durante el mismo se producen intercambios de alelos entre los cromosomas de la pareja de homólogos. Este suceso se denomina sobrecruzamiento o entrecruzamiento y se pone de manifiesto citológicamente por la aparición, entre las cromátidas de los bivalentes, de puntos de cruce, en forma de «X», que se denominan quiasmas. Como sabemos, en cada uno de los dos cromosomas homólogos se encuentran los mismos genes en los mismos loci. Por ejemplo, en el cromosoma número cuatro de la planta del guisante, se hallan, entre otros, el gen responsable de la longitud del tallo (A) y el gen que determina la forma de la legumbre (B). Esta información está en ambos homólogos del par 4, aunque no necesariamente se encuentran en ambos los mismos alelos. En uno de los homólogos puede estar el alelo responsable del tallo alto (A) y el de la forma comprimida de la legumbre (b), mientras que en el otro se pueden hallar los alelos que determinan el tallo corto (a) y la forma de la legumbre hinchada (8) (Fig. 2.9-1). El sobrecruzamiento hace que en los loci de uno y otro cromosoma homólogo aparezcan, tras este proceso, alelos diferentes de los originales, como resultado del intercambio subsecuente al entrecruzamiento. A este proceso, consistente en la combinación de los alelos de los cromosomas homólogos, se le denomina recombinación génica.
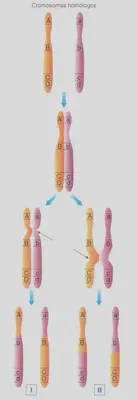
La consecuencia de la recombinación génica es la aparición, en un mismo cromosoma del gameto, de alelos que o bien proceden del padre, o bien de la madre de quien ha producido el gameto. La gran importancia de la recombinación génica es la variabilidad que genera. El número de gametos distintos que se pueden formar mediante este proceso está en función de cuantos loci heterocigotos existen en un individuo. Concretamente, esa cantidad se obtiene elevando el número 2 (par de homólogos) a la cifra de esos loci heterocigotos. Por ejemplo, si un organismo presenta dos loci en heterocigosis, se podrán formar 2² gametos distintos. En nuestra especie se estima que en cada persona existen unos 3.350 loci en heterocigosis. Esto quiere decir que cada individuo puede formar 2 3350 gametos distintos, un número superior al de átomos existentes en el universo. Ello nos da una de las claves del porqué de nuestra individualidad biológica, ya que es prácticamente imposible (salvo en el caso de los gemelos monocigóticos) que existan, hayan existido, o vayan a existir dos personas iguales.
No siempre es posible efectuar intercambios entre los loci de los cromosomas homólogos mediante el sobrecruzamiento. Dado que éste se produce a través de quiasmas, cuanto más juntos estén dos loci, menos probabilidad habrá de que exista sobrecruzamiento entre ellos por un impedimento meramente físico (Fig. 2.9-II). Cuando dos genes tienen nula o muy baja tasa de recombinación entre ellos, se dice que existe ligamiento entre esos dos genes o, simplemente, que están ligados. Si ocurre esto, no existe combinación independiente de caracteres y, por tanto, la ley de la combinación independiente de Mendel queda enmascarada. El porcentaje de recombinación entre dos loci está directamente relacionado con la distancia física que los separa dentro del cromosoma. A más distancia, más porcentaje de recombinación, y viceversa, cuanto más juntos estén menos porcentaje de recombinación se llevará a cabo entre ellos.