Dónde están y qué son los genes: el cromosoma eucariótico y la naturaleza del material hereditario
El descubrimiento del ligamiento y la recombinación propios de la meiosis permitió a principios del sentado un gran avance para este proceso. No obstante, sólo se conocen hasta la fecha un 10-15% de los alrededor de 20.000 genes que se piensa poseemos. En la Fig. 2.10 se ofrece un esquema de la estructura del cromosoma. En el caso de la especie humana, cada célula diploide posee 46 cromosomas; además, sabemos que al tratarse de una especie diploide, hay dos copias de cada cromosoma, por lo que en realidad sólo hay 23 cromosomas diferentes (24 en el caso de los varones, puesto que en los mamíferos el sexo está determinada por la presencia en de un cromosoma especial, el cromosoma Y: en realidad este cromosoma Y hace las funciones de homólogo del cromosoma X, mientras que en las hembras sí existen dos cromosomas X homólogos).
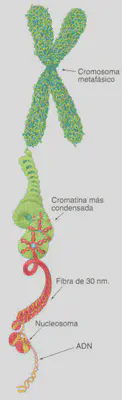
Estructura del Cromosoma Eucariótico
Esencialmente, visto desde el punto de vista de la genética, un cromosoma es una molécula gigantesca de Ácido Desoxirribonucleico (ADN). El ADN es la mayor de las moléculas que portan los seres vivos. Estructuralmente se puede describir como una doble cadena larguísima cuyos eslabones son unas unidades moleculares conocidas como nucleótidos cuyo elemento diferenciador es la base nitrogenada, púrica o pirimidínica, que define a cada uno de ellos. Como luego veremos, cada uno de los eslabones de una de las cadenas va unido por una forma de complementariedad al de la otra cadena y por eso se dice que el ADN está formado por pares de bases (ver Naturaleza Química del Material Hereditario). La longitud de todo el ADN que se encuentra en el total de las células del cuerpo humano es de alrededor de 2 x 10 km. En una célula el tamaño del ADN es bastante menor pero, con todo, alcanza los 2 m. Aunque la delgadez del ADN es extremada, si tenemos en cuenta que el diámetro del núcleo celular es del orden de unas pocas micras, el ADN, dada su longitud, debe experimentar un importante empaquetamiento o condensación, sobre todo en el momento de la división celular, donde esos 2 m de longitud del ADN se convierten en 200 µm en el conjunto de cromosomas metafásicos, es decir, una extensión diez mil veces menor.
Cada cromosoma está constituido por una sola molécula de ácido desoxirribonucleico (ADN) unido a proteínas. Éstas son de varios tipos, pero las principales pertenecen a la familia de las denominadas histonas, que son pequeñas proteínas de carácter básico cuya misión es permitir que el ADN se condense de una forma ordenada alcanzando los diferentes niveles de organización. En eucariotas, el aspecto del material hereditario varía desde la estructura claramente definida que representa el cromosoma metafásico, a una estructura amorfa y disgregada durante la interfase celular, que recibe el nombre de cromatina. Ambas estructuras representan distintos aspectos del mismo material hereditario y ponen de manifiesto que el ADN tiene diferentes niveles de organización. Lo que distingue un nivel y otro es el grado de condensación del ADN. El nivel de organización más elemental (dejando a un lado el que representa la propia molécula de ADN) es el que se alcanza a través de la unión de varios tipos de histonas con el ADN. Esta unión da lugar a una estructura denominada nucleosoma que representa la unidad básica de condensación del ADN (Fig. 2.11). Estas diferencias en el grado de condensación están directamente relacionadas con la funcionalidad del ADN: para que la información contenida en el ADN, lo que llamamos genes, se exprese en la célula es preciso que la molécula sea accesible, por lo que ha de estar poco o nada condensada, mientras que cuando hace falta traspasar esa misma información genética a las células hijas (mitosis) o a los gametos (meiosis), el ADN ha de estar empaquetado, muy condensado. En este segundo caso, la información genética no puede expresarse.
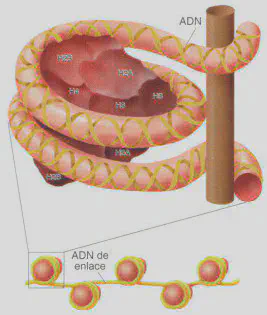
En los eucariotas, el estado menos condensado del ADN consiste en una estructura formada por nucleosomas distribuidos más o menos periódicamente a lo largo del material hereditario como si de cuentas de un rosario se tratase (Fig. 2.11). Esta disposición hace que el ADN disminuya aproximadamente siete veces su longitud. Sin embargo, en el cromosoma metafásico el nivel de condensación del ADN es todavía unas 1.400 veces mayor que el alcanzado sólo con las «cuentas » que forman los nucleosomas. Ese mayor grado de compactación se consigue por sucesivos procesos de plegamiento de unos niveles de organización para alcanzar otros superiores. El nucleosoma representa el primer nivel y el cromosoma metafásico el último. Los sucesivos niveles se consiguen gracias a que los nucleosomas, una vez formados, se pliegan unos sobre otros de una manera ordenada formando una fibra de 30 nm de espesor (Fig. 2.12). Esta fibra representa el siguiente nivel de organización del material hereditario y proporciona una compactación del ADN que hace que su longitud sea 100 veces menor. Los siguientes niveles de condensación no son bien conocidos. En cualquier caso, posteriores enrollamientos de estas fibras de 30 nm, posiblemente sobre un eje proteico no histónico, proporcionan el grado final de condensación del ADN (Fig. 2.10).
Como se ha indicado, la condensación del ADN a lo largo del ciclo celular varía desde el estado de cromatina al de cromosoma metafásico (Fig. 2.10). Sin embargo, la cromatina tampoco presenta un estado homogéneo de compactación y se distinguen, a este respecto, dos tipos de cromatina, la eucromatina, que presenta un empaquetamiento menor, y la heterocromatina, que es la porción de cromatina más condensada. Como veremos a la hora de hablar sobre la regulación de la expresión génica, los diferentes niveles de organización de la cromatina están relacionados también con el grado de expresión génica.

Naturaleza Química del Material Hereditario
Antes de identificar a los cromosomas como los vehículos de la herencia a principios del siglo XX, ya se conocía que estaban formados por proteínas y ácidos nucleicos. Una de las siguientes preguntas era ¿cuál es la naturaleza química del gen? Aun sin conocer la respuesta a esta incógnita, sí se sabía que las propiedades que debía cumplir el material encargado de portar la herencia biológica eran principalmente dos: 1) guardar información y 2) permitir copiar fielmente dicha información. Además, sus propiedades tenían que explicar la cierta capacidad de cambio o de alteración de la propia materia hereditaria, que daría cuenta de la existencia de variantes genéticas o alelos. Durante un tiempo las proteínas, dado el gran número de combinaciones que es posible realizar con los diferentes aminoácidos que pueden formar parte de su estructura, fueron consideradas las candidatas idóneas para ser las moléculas de la herencia. Sin embargo, fue el ácido desoxirribonucleico o ADN, aislado por primera vez en 1869 por el suizo F. Miescher (1844-1895), la molécula que resultó poseer todas las características necesarias. Fueron J.D. Watson (1928-) y F. Crick (1916-2004) los que describieron por primera vez la estructura de la molécula de ADN en el artículo publicado en 1953 en la revista Nature, «Estructura molecular de los ácidos nucleicos: una estructura para el ácido nucleico de la desoxirribosa». Gracias a dicha estructura se pueden explicar las bases moleculares de la herencia biológica. Este tipo de estudios contribuyó definitivamente al desarrollo de la Genética molecular, una disciplina entre cuyos cometidos se encuentra establecer de una forma completa la relación entre genotipo y fenotipo.
Igual que otros ácidos nucleicos, el ADN es una larga cadena doble formada por nucleótidos (Fig. 2.13) que son sustancias compuestas por una molécula de ácido fosfórico, una de un hidrato de carbono -que puede ser una ribosa o una desoxirribosa- y otra de una base nitrogenada -que puede ser una base púrica o una base pirimidínica. En el ADN estos nucleótidos forman dos cadenas, cada una de las cuales está dispuesta en espiral, enroscada una sobre otra formando una doble hélice (Fig. 2.14). Esta conformación se consigue gracias a una disposición con creta de las moléculas que forman cada nucleótido del ADN. La espiral la marca la sucesión de las moléculas de desoxirribosa y ácido fosfórico de cada nucleótido, mientras que las bases nitrogenadas se sitúan en el interior. La unión entre las dos cadenas de nucleótidos que forman el ADN se lleva a cabo a través de puentes de hidrógeno que se establecen entre las bases púricas de una cadena y las pirimidínicas de la otra.
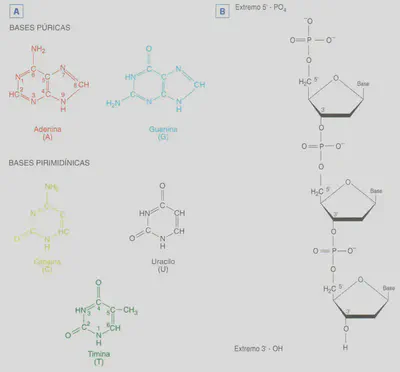
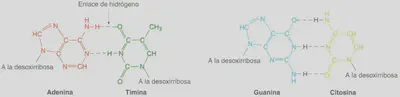
Existen severas restricciones acerca de las uniones entre las bases nitrogenadas de las dos hebras o cadenas que constituyen el ADN, ya que la adenina se aparea únicamente con la timina, mientras que la citosina lo hace sólo con la guanina (Fig. 2.15). A esta relación restrictiva entre las bases se le denomina complementariedad y hace que las dos cadenas de nucleótidos del ADN sean complementarias entre sí. Esta propiedad es la causa de la relación 1:1 entre bases púricas y pirimidínicas. El modelo propuesto por VVatson y Crick explica ese hecho: dado que una base púrica se aparea siempre con la misma base pirimidínica (A - T y C - G), la cantidad de bases púricas será siempre igual a la de pirimidínicas, es decir, A+ G = T + C ó A/ T = C / G. Por ejemplo, si en una determinada molécula de ADN la timina representa e! 17% de todas las bases nitrogenadas de ese ADN, dado que esta base únicamente se aparea con la adenina, la cantidad de ésta también representará el 17% de las bases nitrogenadas de la molécula de ADN analizada. El porcentaje restante, 66%, estará repartido a partes iguales entre la otra pareja de bases complementarias: 33% para la citosina y 33% para la guanina.
Las Copias para la Herencia: Duplicación del ADN
Antes de analizar cómo se codifica la información genética vamos a estudiar cómo se copia el ADN, paso imprescindible para poder traspasar la información a la siguiente generación. La clave está en la complementariedad de las bases nitrogenadas de las dos cadenas que lo forman. Las características fundamentales del proceso de replicación del ADN y los mecanismos mediante los cuales las enzimas encargadas lo llevan a cabo son prácticamente similares en todos los organismos. En 1958, M. Meselson (1930-) y F.W. Stahl (1929-) demostraron que la replicación del ADN es semiconservativa, es decir, que a partir de una molécula de ADN se obtienen dos, cada una de las cuales porta una hebra del ADN original, en tanto que la otra hebra o cadena complementaria se va sintetizando siguiendo la complementariedad original (Fig. 2.16). Cuando el proceso concluye, las dos nuevas moléculas de ADN se separan. Ambas llevan una hebra antigua y otra nueva, pero las dos son idénticas: de esta forma, la información puede ser trasmitida fielmente a otra generación. Cuando hayan de traspasarse a las células hijas o a los gametos, se empaquetarán, unida la una a la otra por el centró mero para constituir el cromosoma metafásico que, como sabemos, contiene dos cromátidas, es decir, dos dobles cadenas resultado de la replicación del ADN.
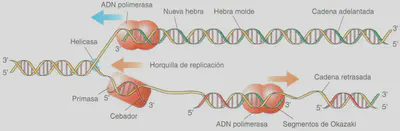
La afinidad sumamente específica entre entra las bases complementarias, que hace que sea enormemente difícil (aunque no imposible, como veremos) que sea una base inapropiada la que se empareje como complementaria en el proceso de replicación, hace del ADN, en efecto, una molécula química idónea para guardar la información genética.